Le longicorne asiatique est un insecte forestier envahissant originaire de Chine et de Corée. Il se nourrit du bois de presque toutes les essences de feuillus en Amérique du Nord, mais spécialement des érables indigènes. Les femelles pondent leurs œufs à travers l’écorce et les larves creusent des tunnels dans les tissus vivants, interrompant ainsi le transport de l’eau et des nutriments et entrainant la mort de l’arbre. L’insecte se transporte facilement dans le de bois de chauffage, d’arbres vivants ou par du bois non traité, comme le matériel d’emballage utilisé pour le transport maritime de marchandises.
Le longicorne asiatique a été introduit pour la première fois en Amérique du Nord dans les années 1990, probablement par des matériaux d’emballage en bois massif, telles des caisses ou des palettes de bois. En 2003, il a été découvert pour la première fois au Canada dans un parc industriel de la région de Toronto. Un programme d’éradication intensif a alors été mis en place pour retirer les arbres hôtes dans une zone de quarantaine autour de l’infestation, au coût de dizaines de millions de dollars sur quatre ans, afin d’empêcher la propagation. Une seconde infestation a été détectée à Toronto en 2013. Des mesures de détection et d’éradication ont, de nouveau, été mises en place et un programme de surveillance est présentement en cours pour évaluer le succès de ces efforts. Au total, c’est près de 500 millions de dollars américains qui ont été dépensés en Amérique du Nord depuis les années 1990 pour l’éradication et la surveillance du longicorne asiatique.
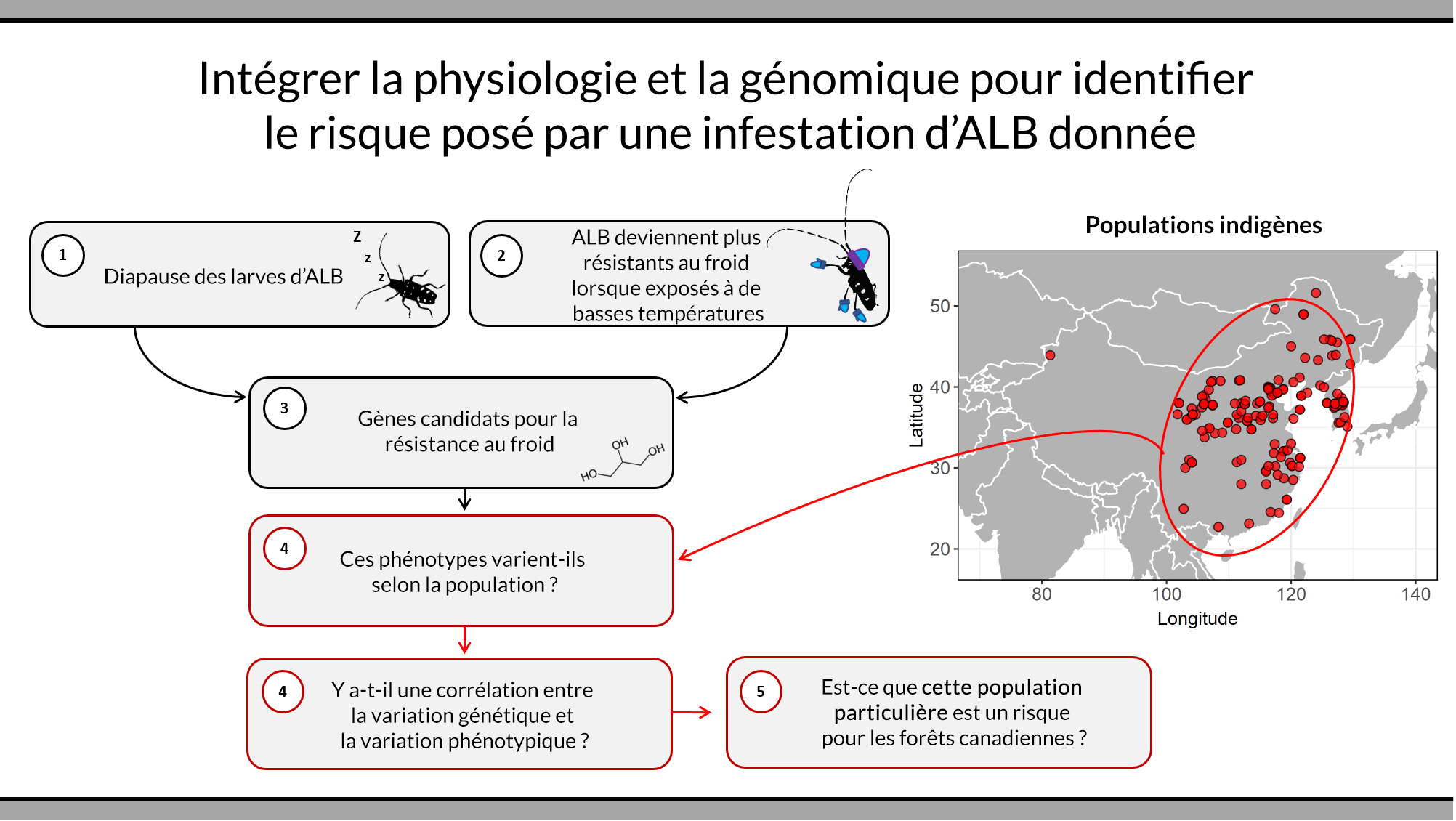
Les populations de longicorne asiatique colonisent de nombreux milieux en Chine et nous nous attendons à ce que certaines d’entre elles soient plus résistantes aux rudes hivers canadiens. Si nous pouvons identifier ces populations, les ressources pourront être allouées aux introductions les plus à risque. Une partie de l’équipe de bioSAFE (dirigée par Dre Ilga Porth de l’Université Laval et Dre Amanda Roe de Ressources naturelles Canada) identifie des marqueurs génétiques appelés polymorphismes nucléotidiques (SNP en anglais) qui nous permettent d’identifier efficacement l’origine géographique d’une l’infestation.
Une approche parallèle et connexe est dirigée par Dr Brent Sinclair (Western University), Dre Amanda Roe et Dr Daniel Doucet (Ressources naturelles Canada) et tente d’identifier des marqueurs génomiques (figures 1 et 2) indiquant la capacité d’un individu de longicorne asiatique à survivre à l’hiver. Pour ce faire, ils déterminent d’abord les mécanismes physiologiques responsables de la résistance au froid puis utilisent la transcriptomique et la métabolomique pour identifier les cascades de réactions et les molécules qui sous-tendent ces mécanismes. Ils utiliseront ensuite la base de données de SNPs pour identifier ceux qui sont susceptibles d’être spécifiquement associés à une capacité d’hivernage accrue. Ces travaux sont effectués au Laboratoire de quarantaine et de production d’insectes du Canada, la seule installation canadienne certifiée pour l’hébergement de colonies de longicorne asiatique.
Figure 1.
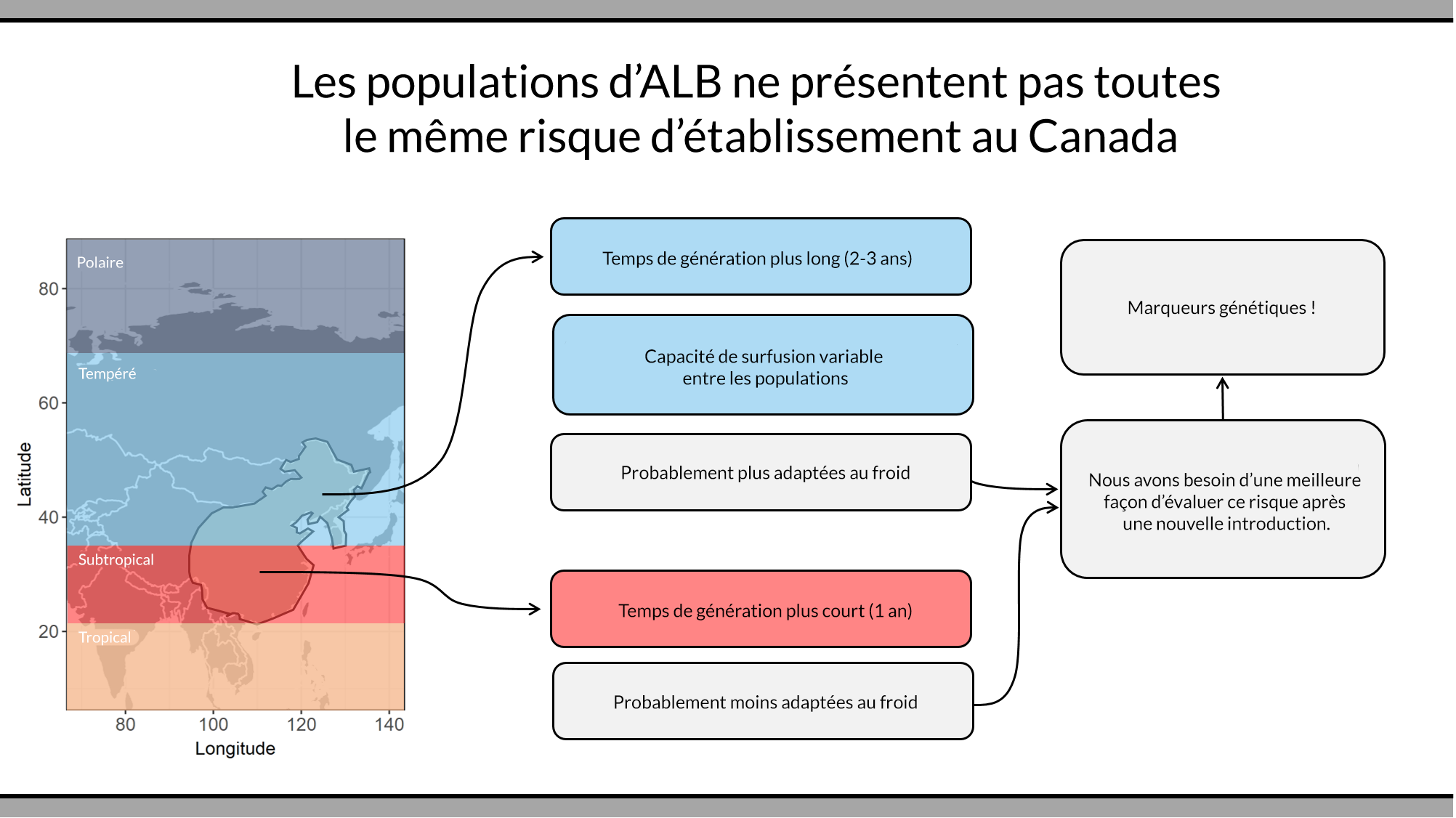
Figure 2
Organigrammes originaux par Dr. Alex Torson